本系列将更新三维基因组相关内容,欢迎关注!
二维顺式元件
术语 cis 源自拉丁语词根“cis”,意思是“同一侧”。相比之下,“trans”一词来自拉丁语词根“trans”,意思是“对面”。在分子生物学中,顺式调控元件是指染色体 DNA 中调节同一染色体上基因转录或表达的区域。反式作用因子是指与基因的顺式作用元件结合以控制其表达的可溶性蛋白质。然而,可溶性反式作用蛋白可以驻留在任何染色体上,通常位于与其调节位置不同的染色体上。
生化活性调控元件(由序列特异性调控转录因子结合):
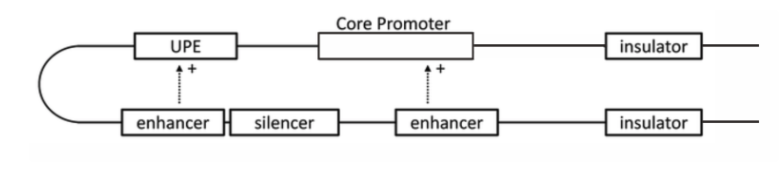
- 启动子:启动子是基因 TSS (+1) 周围的区域,其中包含多个有助于调节蛋白结合的 DNA 元件。它为 RNA 聚合酶和招募 RNA 聚合酶以使转录发生的蛋白质(转录因子)提供安全的初始结合位点。
- 增强子:增强子是CRE(顺式调节元件,这意味着它们是非编码DNA,不编码转录因子但参与调节)。它们距离基因最远可达 1M bp (1,000,000 bp),位于起始位点的上游或下游。
- 绝缘子:绝缘子是阻止增强子和启动子之间相互作用的遗传边界元件。人们发现它聚集在拓扑关联域(TAD)的边界,并且可能在将基因组划分为“染色体邻域”(发生调控的基因组区域)方面发挥作用。
- 沉默子:沉默子是能够结合转录调控因子的DNA序列,阻止RNA聚合酶与DNA序列的结合,从而阻止基因表达为蛋白质。
多尺度折叠
最大的染色体包含数亿个碱基对,这些碱基对在有限的空间中折叠,从而产生多尺度的分层结构,例如:核小体、染色质纤维、染色体结构域、区室,最后在染色体区域中。
信息存在于各个层面,从亚核小体尺度的组蛋白-DNA相互作用到核空间中的染色体-染色体和染色体-核纤层相互作用。这种多级结构可以通过转录因子、结构蛋白和非编码RNA等多种成分来调节和/或利用,以协调基因表达和细胞命运。
借助当前开发的染色体捕获技术,我们可以看到它们如何扩展我们对染色体结构的了解。

染色质疆域
在更大的尺度上,染色质被组织成单独的染色体区域(每条染色体一个),这些区域很少混合。这一观察结果最初来自 FISH 研究,后来通过全基因组 Hi-C 数据进行了验证,该数据表明同一染色体上的基因座之间的相互作用比不同染色体之间的反式接触要频繁得多。
A/B Compartments
Hi-C 数据经过归一化并转换为观察/预期矩阵后,显示格子图案,通过主成分分析 (PCA) 进行分析,第一个主成分(可以解释为数据最大方差的替代项,最突出的特征)包含区室 A 和 B(阳性 PC1 区域反映“活跃/允许”染色质,阴性 PC1 区域表示“非活跃/惰性”染色质)。

TAD 拓扑结构域
拓扑关联域 (TAD) 是一个自相互作用的基因组区域,这意味着 TAD 内的 DNA 序列之间的物理相互作用比 TAD 外的序列更频繁。这些三维染色体结构存在于动物以及一些植物、真菌和细菌中。 TAD 的大小范围从数千到数百万个 DNA 碱基不等。
TAD 通常表现为沿 Hi-C 图对角线的连续方形域。基因组到 TAD 的空间划分与许多线性基因组特征相关,例如组蛋白修饰、协调基因表达、与核纤层和 DNA 复制时间的关联、增强子-启动子相互作用。
TAD 的边界富集:
- 绝缘体蛋白:CTCF(在所有边界的约 76% 处检测到)
- 活性转录标记:H3K4me3 和 H3K36me3
- 新生转录本
- 看家基因(存在于大约 34% 的 TAD 边界)
- 重复元件
还有证据支持 TAD 在不同细胞类型和物种之间是保守的。
- TAD 的定位在不同细胞类型中相对稳定,并且似乎独立于组织特异性基因表达或组蛋白修饰。在 ESC 分化过程中,区室 A 和 B 发生全基因组切换,而 TAD 定位保持稳定 。
- TAD 定位在进化上是保守的:50-70% 的 TAD 边界在人类和小鼠 ESC 之间共享 。
- TAD 是复制时间调节的稳定单位。

Sub TAD
TAD 可以进一步分为从小鼠 ESC 的高分辨率 5C 观察到的更小的子 TAD 。它类似于 TAD 显示自关联特征,跨子 TAD 边界的接触频率降低,并且一些子 TAD 边界与 CTCF/Cohesin 介导的相互作用相关。然而,与 TAD 不同的是,亚 TAD 在细胞/组织类型中不太保守,并且似乎与细胞类型特异性基因表达相关。
染色质环
人们已经认识到,像启动子-增强子这样的顺式调控元件通常沿着脊椎动物的线性基因组远离。然而,为了引发调节作用,基因组结构进化形成一个环,将两个元素聚集到空间上接近的位置。这种染色质形成通常称为“染色质环”。一个众所周知的例子是 β-珠蛋白簇的基因座控制区 (LCR),它通过长距离染色质接触与其在类红细胞中的靶基因(其中 β-珠蛋白基因活跃)强烈相互作用,但不同谱系的细胞之间几乎没有相互作用。
核小体-核小体相互作用
这是染色质组织的最小规模。长期以来,基于体外电子显微镜,核小体被认为形成具有螺线管或之字形形状的阵列(通常称为 30 nm 染色质纤维)。然而,最近的研究提供了更多证据来支持更灵活、异质的群体排列结构。
结构蛋白和 RNA
染色质生物学中的一个重要问题是如何建立 3D 染色质组织的结构特征。很少有结构蛋白被证明对染色质结构至关重要。
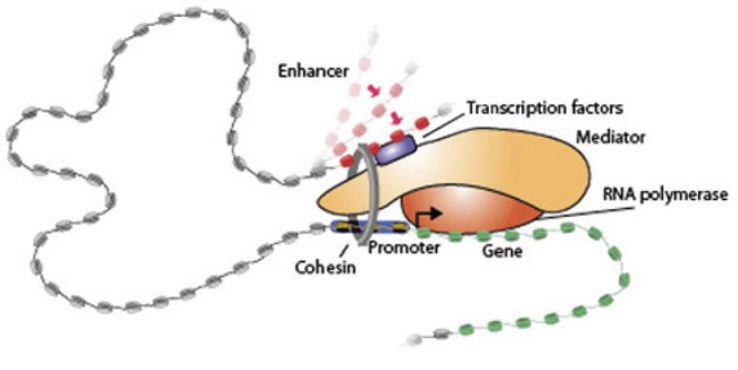
介体(共激活剂)
介体存在于活跃转录基因的增强子和启动子中,并通过启动前复合物 (PIC) 组装和 RNAPII 延伸来促进转录。
粘连蛋白
粘连蛋白是具有多种功能的蛋白质:
- 它在细胞分裂过程中调节姐妹染色单体的分离。
- 它对于DNA修复很重要。
- 对于染色质结构:粘连蛋白与 CTCF 和介体相互作用被认为是间期细胞中环挤压复合体的一部分。
CTCF
CTCF 最初被定性为绝缘体蛋白,能够限制增强子-启动子相互作用。大约 15% 的 CTCF 在哺乳动物的 TAD 边界处富集,大多数位于 TAD 内,并且被认为参与了 TAD 内的相互作用。另一个显着特征是环锚定处的 CTCF 位点主要以会聚方向出现,这表明不仅结合而且结合序列的方向性对于环的形成也很重要。
非编码RNA结合
一个有趣的观察是介体和 CTCF 似乎都能够直接与 RNA 结合,并且敲除一些介体结合非编码 RNA 导致 ncRNA 位点与其靶标之间的环形成减少。
未完待续,持续更新,欢迎关注!








 苏公网安备32011502012024号
苏公网安备32011502012024号